Без генетске варијабилности, сва жива бића би (наслеђивањем) требала бити иста као прва. За неједнака бића једино би објашњење било оно које се односи на појединачне креације. Али знамо да структура ДНК, која је основа за пренос наследних карактера, има релативну, а не апсолутну стабилност. Док стабилност гарантује очување почетних информација, нестабилност одређује модификације, или боље (да употребимо одређени израз) мутације.
Мутације се могу поделити у 3 велике групе:
- мутације гена;
- хромозомске мутације;
- геномске мутације.
На овом месту је добро укратко додати два концепта: један је онај о „поправци“, а други о „повољној“ или „неповољној“ мутацији.
Поправак, енглески израз који значи поправка, односи се на постојање у ћелијама специфичних ензимских процеса који имају за циљ да провере очување информација, елиминишући или исправљајући делове ДНК који нису у складу са оригиналом.
Концепт "повољне" или "неповољне" мутације изражава поређење између ефикасности, за организам и за врсту, оригиналног гена ("дивљи", тј. "Дивљи", примитиван) и мутираног гена.
Треба напоменути да се предности и недостаци увек односе на одређене услове животне средине; неповољна мутација у одређеном окружењу може постати повољна под различитим условима.
Последице овог принципа су огромне у популационој генетици, што се већ може показати различитом ефикасношћу преживљавања варијанти хемоглобина у нормалном или маларичном окружењу. Али ову тему треба одложити на детаљније чланке о општој, хуманој или медицинској генетици.
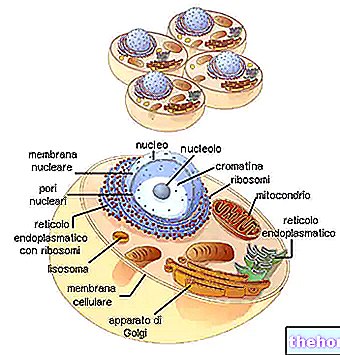
Потребна је још једна разлика (посебно за мутације гена) између соматских мутација и мутација заметне линије. Све ћелије нашег вишећелијског организма потичу, као што је познато, из једне зиготе, али се врло рано диференцирају у соматску линију, из које се развија цело тело, и заметну линију предодређену да формира гонаде, а за терминалну мејозу, гамете Сасвим је евидентно да ће се, осим интеракција између две ћелијске популације, мутација соматске линије манифестовати у једном организму, али неће утицати на потомке, док ће се мутација у заметној линији појавити само у потомцима.
С обзиром на различите типове мутација и релативне последице, вреди поново споменути класификацију узрока мутације. Ови узроци се називају мутагени, који се користе за разликовање углавном на физичке и хемијске мутагене. Разне модификације физичког окружења могу довести до мутација, али главни физички мутагени су зрачење.Због тога су радиоактивне супстанце опасне, а прије свега теже радиоактивне честице изазивају мутације усљед дефекције, са најтежим посљедицама.
Хемијски мутагени могу деловати и променом уређене структуре нуклеинских киселина, и уносом у ћелије супстанци сличних нормалним азотним базама, које могу да се такмиче са последњим у синтези нуклеинских киселина, изазивајући тако мутације супституцијом.
ГЕНСКЕ МУТАЦИЈЕ
Генске мутације утичу на један или неколико гена, односно на ограничени део ДНК. Пошто се информације чувају у низу нуклеотидних парова, најмања јединица мутације (мутон) састоји се од једног комплементарног пара база. Без улажења у детаље о различитим механизмима мутације на нивоу гена, можемо се ограничити на помињање два: супституција базе и поновни избор (или уметање). У мутацијама супституционе базе, један или више нуклеотида ДНК се замењују другим. Ако грешка није исправљена на време, следи низ у време транскрипције такође промењене у РНК. Ако је промена триплет није ограничен на синоним (види генетски код), то ће резултирати супституцијом једне или више аминокиселина у полипептидној секвенци. Замена аминокиселине може бити мање или више критична за одређивање структуре протеина и за његову функцију.
Код поновних избора или инсерционих мутација, један или више нуклеотида се уклања или додаје у ДНК секвенцу. Ове мутације су генерално веома озбиљне јер се (осим ако се ради о целим тројкама које додају или одузимају појединачне аминокиселине) мењају све триплете које следе по редоследу читања.
Генске мутације су најчешће и извор су већине варијабилности наследних особина међу појединцима.
ХРОМОСОМСКЕ МУТАЦИЈЕ
Ово су мутације које утичу на релативно дугачке фрагменте хромозома. Уобичајено их је класификовати углавном на:
- хромозомске мутације за реизбор;
- хромозомске мутације дупликацијом;
- хромозомске мутације транслокацијом.
Мутације услед дефекције настају услед лома и губитка мање или више дугачког фрагмента хромозома. Посебно у мејози, ова врста мутације је често смртоносна, због потпуног губитка одређеног броја гена, више или мање неопходних.
У дупликационим мутацијама, након паузе, хроматидни пањеви имају тенденцију да се споје.
У накнадном уклањању центромера, хромозом, који је постао дицентричан, ломи се на често неједнаке делове: као што видимо, резултат је реизбор с једне стране и дуплирање с друге стране.
Хромозомски прекид може бити праћен преокретом. Укупан генетски материјал је непромењен, али је секвенца гена на хромозому промењена.
Случај транслокације је сличан, али се односи на везивање фрагмента хромозома за нехомологни хромозом. Један хромозом је ампутиран, а други продужен; укупна генетска информација ћелије је и даље непромењена, али је ефекат положаја још израженији. Лако је представити ефекат позиције позивањем на концепт регулације деловања гена: променом положај на хромозомима, ген може лако напустити један оперон и бити уметнут у други, што доводи до „измењене активације или потискивања.
Међутим, каже се да је транслокација уравнотежена (или уравнотежена) када постоји реципроцитет транслокације између два пара хромозома, при чему збир информација о гену остаје непромењен. Фигура у облику крста у мејотичкој дијакинези генерално одговара уравнотеженој транслокацији.
ГЕНОМСКА МУТАЦИЈА
Имајући у виду да је геном индивидуална генетска баштина, поређана у хромозомима, може се специфицирати да говоримо о геномским мутацијама када хромозоми имају другачију дистрибуцију од норме врсте.
Геномске мутације се могу класификовати углавном у мутације због полиплоидије, хаплоидије и анеуплоидије.
Полиплоидне мутације се јављају када редупликација не резултира поделом; лакше се јављају у биљкама, где се заиста користе за побољшање производње.
Ако ћелијска деоба не успе у мејози, могу се појавити диплоидне гамете; ако таква гамета успе да се споји са нормалном гаметом, зигота која настане овим оплођењем биће триплоидна. Таква зигота понекад може успети у стварању читавог организма, будући да редупликација и митоза не захтевају паран број хромозома. У време мејозе, међутим, редовно упаривање хомологних хромозома биће немогуће.
Геномске мутације хаплоидијем могу настати када се у нормално диплоидној врсти гамета активира другом гаметом без нуклеарног материјала, или чак у одсуству оплодње: резултат ће бити хаплоидна јединка.
Док се претходне врсте геномских мутација увек тичу сабирања или одузимања целог броја н хромозома, мутације за анеуплоидију тичу се вишка или дефекта појединачних хромозома (хромозомске аберације).
Еуплоидни хромозомски састав врсте дефинише се као њен нормални кариотип или идиотип.
















.jpg)

