БЕЛЕШКЕ О ЕМБРИОЛОГИЈИ
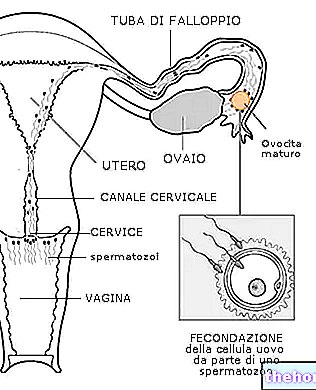
Ембриологија проучава редослед облика развоја од зиготе до организма обдареног свим њеним органима и системима.
С тим у вези, вреди се сетити разлике између развоја (сукцесије структурних и организационих фаза са све већом сложеношћу) и раста, намењеног пре свега у квантитативном смислу.
У метазоанима кичмењака свједоци смо пораста у еволуцијском низу све до човјека (преко циклостома, риба, водоземаца, гмазова, птица и сисара), појављивања одраслих облика све веће сложености, због којих долази до компликација у фазама ембрионалног развоја.
На почетку, зигота, увек опремљена резервним материјалом, подељена је (накнадном митозом) на 2, затим 4, затим 8 итд. Ћелије зване бластомере, без раста, све док не достигне нормалан однос језгра / цитоплазме врсте .
Ова почетна сегментација може пратити различите обрасце, у зависности од количине и дистрибуције деутоплазме.
У почетку је деутоплазма оскудна („олиголецитна јаја“), па је сегментација потпуна и доводи до веома различитих бластомера. Како се повећава сложеност ембриона, потребно је више времена и материјала пре него што му развој дозволи да почне. самосталан живот. Ово захтева повећање деутоплазме ("телоцитна јаја"), која тежи да буде распоређена у делу зиготе. Ово узрокује све већу „анизотропију“, која је повезана са модификацијама сегментације, којима управљају два општа принципа:
- Хертвигов закон каже да, у митози, ахроматско вретено (чији екватор одређује равнину деобе ћелија ћерки) тежи да буде распоређено у правцу највеће дужине цитоплазме;
- Балфуров закон каже да је брзина сегментације обрнуто пропорционална количини деутоплазме.
Тада видимо да је већ код циклостома и код риба сегментација неједнака, са брзо сегментираним животињским полу (који ће дати горње структуре ембриона) и жуманцетом који ће садржавати већину резервног материјала. Ова тенденција је још већа анизотропни код водоземаца (у којима је потребно припремити органе одговорне за дисање ваздухом), у којима пол жуманцета, док се полако сегментира, остаје релативно инертан и завршава покривен ћелијама изведеним из брзо сегментираног животињског пола. главне ембрионалне фазе укључују: зиготу, бластомере, морулу (скуп бластомера сличних купини), бластулу (морулу са регресираним унутрашњим ћелијама), гаструлу (бластулу у којој су ћелије једне стране постале инвагиниране), у којој је примитивна шупљина организма, са спољним ћелијским слојем (ектодерм, из којег ће пре свега проистећи нервни систем) и инт ерно (ентодерм), између којих ће се затим умешати трећи слој (мезодерм). Из ових слојева или "ембрионалних плоча" тада ће, у одређеном редоследу, бити изведена сва ткива, органи и системи.
Код још развијенијих врста повећање деутоплазме (или "телета") је такво да се не може ни сегментирати. Тако видимо да сегментација код птица утиче само на танки површински диск, што доводи до "дискобластуле" и низа појаве које гарантују формирање ембриона на другачији начин од горе поменутог.
Даље повећање деутоплазме вероватно не би било ефикасније, па се код сисара развој и раст до способности за самосталан живот постижу другим системом. У ствари код сисара примећујемо да се деутоплазма користи само у првим фазама развој; тада ембрион успоставља метаболичке односе са мајчиним организмом (са појавом плаценте) и више не користи деутоплазму, чији се вишак елиминише. У овом тренутку јаја се враћају у олиголектит и сегментација се може вратити да је тотална (и стога је у раним фазама слична оној код "амфиоксуса"), али након моруле ембриогенеза се наставља према најразвијенијем обрасцу птица, са "бластоцистом" након чега следи имплантација на зид материце, тако да метаболизам ембриона обезбеђује мајчински организам (преко плаценте), а не деутоплазма.
ЕМБРИЈСКА ДИФЕРЕНЦИЈАЦИЈА
Када је сегментацијом зиготе однос језгра / цитоплазме доведен у норму врсте, раст такође мора започети, паралелно са развојем. Из тог разлога почиње метаболизам, са појавом нуклеола и синтезом протеина. Тако започета синтеза протеина је последица гена одговорних за ране фазе ембрионалног развоја. Ови гени су потиснути супстанцама присутним у различитим бластомерима животињског и телећег стуба. Заузврат, производи ових раних гена могу смањити притисак на опероне гена задужених за касније фазе. Производи ове друге серије гена моћи ће да делују како у смислу конструисања нових ембрионалних структура, тако и у смислу потискивања претходних оперона и дерепресије следећих, у уређеном низу који доводи до изградње новог организма , захваљујући генетичким информацијама прикупљеним из генома кроз миленијуме у све развијеније врсте.
Хаецкелов чувени израз "онтогенеза резимира филогенију" заправо изражава чињеницу да више врсте понављају, у фазама ембрионалног развоја, сукцесију која је већ пронађена у еволуцијски претходним врстама.
Ране фазе ембриона имају тенденцију да буду сличне код кичмењака, посебно до појаве шкрга.
Код врста које прелазе на ваздушно дисање, шкрге се затим поново апсорбују и поново користе (на пример за формирање ендокриних жлезда), али су генетски подаци који се односе на стварање шкрга сачувани и код људи. Ово је очигледно пример ембрионалних структурних гена који су присутни у геному свих кичмењака и морају остати потиснути након што су функционисали у свом онтогенетском тренутку.
Тумачење ембриогенезе у смислу регулације деловања гена омогућава обједињавање сложених традиционалних искустава експерименталне ембриологије.
БЛИЗАНЦИ
Жигота и први бластомери, све док не почне синтеза протеина, су тотипотентни, односно способни да оживе читав организам. С тим су повезани експерименти Спеманна, који је добио два ембриона из дављења зиготе водоземаца. Сличан феномен јавља се у основи феномена једнојајчаних близанаца код човека, који се управо из тог разлога називају монозиготним (МЗ) .Спеманнови експериментални близанци били су упола мањи од нормалног, док су код човека савршено нормални. То се објашњава јер су код водоземаца два ембриона морала да деле једино жуманце које су већ примили, док су код човека ембриони могли да приме, преко плаценте, све што је потребно за њихов развој и раст.
Треба запамтити да код „човека две трећине случајева близанаца“ има друго порекло: они потичу од повременог истовременог сазревања два фоликула, са ослобађањем два овала који, када су оплођени, дају две зиготе; у ствари, у у овом случају говоримо о двојајчаним близанцима (ДЗ).
Будући да близанци МЗ, подељени митозом од појединачне зиготе, имају исти геном, разлике између њих морају бити еколошког порекла. Уместо тога, геном два близанца ДЗ сличан је само онолико колико и код било које друге браће. Метода близанаца, нашироко користи у хуманој генетици и у области спорта.
У „човеку, у коме би одређени етички разлози забрањивали експериментисање, може се утврдити колико је неки карактер регулисан наследним факторима: у ствари, строго наслеђени карактери (попут крвних група) увек су подударни само код близанаца МЗ; подударност особине у МЗ блиска је оној у ДЗ, закључује се да фактори средине преовлађују над наследним у одређивању те фенотипске особине.